На основании этого вводится показатель q - коэффициент генеративной емкости жизненного цикла:

где: Т - общая продолжительность жизненного цикла рыбы;
t - возраст созревания рыбы.
На основании анализа, проведенного автором значение данного коэффициента для исследованных рыб оказались выше 0,5(q≥0,5), что объясняется естественным эволюционным отбором наиболее рационально организованных репродуктивных структур, обеспечивающих быстрое воспроизводство и наращивание численности. При этом зависимость годовой скорости воспроизводства от возраста наступления половой зрелости выражается степенным уравнением регрессии вида:

где: значения коэффициентов a = 1,596 и b = - 0,136 получены на основе проанализированных популяций рыб. Построенный по данной формуле график (в виде гиперболы) характеризует, как резко снижается репродуктивный потенциал популяций по мере увеличения сроков их созревания.
Данный принцип (зависимость) учитывается при определении величины ПДУ, то есть, чем меньше возраст полового созревания, тем выше должен быть допустимый улов и наоборот.
110. Согласно предлагаемого принципа, основанного на концепции репродуктивной популяции, учитывая возраст массового наступления половозрелости, определены максимально возможные значения коэффициента промысловой смертности для каждого вида рыб (таблица 26).
Таблица 26. Значения коэффициентов промысловой смертности рыб
| Виды рыб | Возраст созревания | Коэффициент промысловой смертности φF |
| лещ | 4 | 0,266 |
| судак | 4 | 0,266 |
| сом | 5 | 0,234 |
| сазан | 4 | 0,266 |
| жерех | 4 | 0,266 |
| берш | 4 | 0,266 |
| вобла | 4 | 0,266 |
| карась | 4 | 0,266 |
Принимая во внимание полученные значения коэффициентов естественной и промысловой смертности, получается коэффициент общей смертности для каждого вида рыб, исходя из равенства:

Полученное таким образом значение коэффициента общей смертности, дифференцированный по возрастам приведен в столбце № 8 таблицы 25.
Для дальнейшего расчета ПДУ необходимо определить абсолютную численность промысловых видов рыб.
111. Численность рыб определяется методом площадей по результатам неводной съемки по методу ВНИПРХ, с использованием соответствующих работ и методик. Рассчитанные значения абсолютной численности леща озера Балхаш приведены в колонке № 3 таблицы 25. В колонках № 2 и № 6 приводятся данные ихтиологических наблюдений. В колонке № 7 приводится абсолютная величина численности родительского стада для всех возрастов, полученная перемножением данных колонок № 3 № 6. В колонке № 9 дается остаточная численность рыб после года промысла. Например, остаток пятилетних рыб, ставших к концу года шестилетними, равен:
20696 * (1 - 0‚5018) = 10385,25 тысяч штук
В колонке № 10 приводится ихтиомасса остаточной численности рыб. В колонке № 11 приводится численность родительского стада в конце года промысла. В колонке № 12 определяется фактическое значение коэффициентов естественной смертности - φ1m. Например, для шестилетнего леща озера Балхаш:
φ1m = (1 - 0,266) 0,2572 = 0,1888
В колонке № 13 даются фактические значения коэффициентов промысловой смертности (убыли от вылова) - φ1F. Для тех же шестилетних рыб, к примеру:
φ1F = 0,5232 - 0,1888 = 0,3344
В колонке № 14 дана величина улова по возрастам в тысячах штук. Например, для рыб в возрасте шести лет улов равен 9432 * 0,3344 = 3154,2 (тысяч штук), а масса этого улова (колонка № 16) равна 3154,2 (тысяч штук) * 221,75 (грамм) = 699,44 (тонна).
Значения величины средней массы вылавливаемой рыбы (колонка № 15) найдены с учетом допущения, что рыба в течение года вылавливается равномерно, и могут быть выражены формулой:
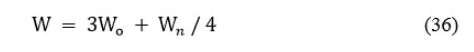
где: Wo и Wn - средняя масса вылавливаемой рыбы в начале и конце года. Например, для шестилетних рыб:
W6 = 3W6 +W7 / 4 = 3 * 210 + 257 / 4 = 221,75 (грамм)
Таким образом, согласно расчетов, приведенных в таблице 25, предельно допустимый улов леща озера Балхаш может составить 4000 тонн.
112. Значения абсолютной численности рыб, ихтиомассы, абсолютной численности родительского стада в конце года, для рационально эксплуатируемой промыслом популяции, должны быть не меньше этих значений в начале года. Но в зависимости от биологического состояния популяции вида, интенсивности промысла в предыдущие годы, целевых ориентиров формирования промысловой ихтиофауны на перспективу эти значения могут отличаться.
В случае, если данные о величине естественной смертности отсутствуют, приближенное значение целевого ориентира по интенсивности промысла можно рассчитать на основе концепции репродуктивной разнокачественности популяций, согласно которой скорость увеличения численности популяций рыб зависит от возраста их созревания и числа повторных генераций в течение жизни.
При этом индивидуальная плодовитость имеет второстепенное значение. Выразив промысловую смертность в терминах коэффициента годовой убыли получают биологически приемлемые значения этого коэффициента для видов, отличающихся возрастом полового созревания самок.
Два принципиальных момента, которые следует учитывать для правильной трактовки оценок годовой убыли:
1) допустимая доля промыслового изъятия выражена в процентах от численности запаса, поэтому и оценка улова будет в единицах численности, для нахождения величины допустимого улова в единицах массы полученную оценку необходимо пересчитать с учетом возрастного состава запаса и средних навесок;
2) допустимая доля промыслового изъятия относится к промысловой части запаса в начале прогнозного года.
Управление рыболовством должно основываться на концепции максимального среднемноголетнего улова (далее - МSY). Коэффициенты изъятия промыслового запаса проверяются на соответствие критерию МSY. В случае несоответствия производится расчет иных коэффициентов изъятия промыслового запаса.
Проверка соответствия критерию MSY осуществляется путем моделирования состояния промыслового запаса и ПДУ на два последовательных года. При этом, критерий MSY состоит в том, что ПДУ на второй год промысла должен быть больше или равен ПДУ на первый год промысла, а объем промзапаса на второй год промысла больше или равен объему промыслового запаса в первый и нулевой годы (пример расчета 1).
1) Для видов, по которым достигнуты граничные ориентиры запаса по биологическим показателям рыб LС50 ≤ LМ50 применяется формула Z=2F, то есть коэффициент изъятия F рассчитывается с коэффициентом 0,5 от коэффициента общей смертности Z, и значения F берутся из таблицы 27.
2) Для популяций, у которых LC50 незначительно больше LM50 (на 1-2 сантиметра, то есть приближается достижение граничных ориентиров запаса), принимается, что Z (коэффициент общей смертности) = M (коэффициент естественной смертности) + F (коэффициент промысловой смертности, он же коэффициент изъятия), а F=M (в соответствии с рекомендациями ФАО для популяций рыб, достигших граничных ориентиров состояния запаса). При этом, сначала F находится по таблице Малкина (таблица) или уравнению Риклефса (пункт 94).
Проверка значения F на соответствие критерию МSY проверяется посредством включения данных полученных по таблице Малкина или уравнения Рифлекса в матрицу расчетов. При несоблюдении критерия (ПДУ и промзапас на второй год промысла снижаются или сильно растут), производится пересчет, изменяя значение F, пока ПДУ2 не будет больше или приблизительно равно ПДУ1.
Пример расчета 1
| Средняя масса, грамм | Nt0, тысяч штук | Bt0, тонна | Половозрелость, доли единиц | Pt1, тонн | F | M | Z | ПДУ 1 года, тонн | Nt+1, тысяч штук | Bt+1, тонна | Pt+1, тонна | ПДУ 2 года, тонна | Nt+2, тысяч штук | Bt+2, тонна | Pt+2, тонна |
| 83 | 113 | 9,379 | 0,5 | 4,6895 | 0,28 | | 0,56 | 1,31306 | 113 | 9,379 | 4,6895 | 1,31306 | 113 | 9,379 | 4,6895 |
| 137 | 106 | 14,522 | 0,6 | 8,7132 | 0,28 | | 0,56 | 2,439696 | 49,72 | 6,81164 | 4,086984 | 1,14435552 | 49,72 | 6,81164 | 4,086984 |
| 206 | 24 | 4,944 | 0,8 | 3,9552 | 0,28 | | 0,56 | 1,107456 | 46,64 | 9,60784 | 7,686272 | 2,15215616 | 21,8768 | 4,5066208 | 3,60529664 |
| 380 | 2 | 0,76 | 1 | 0,76 | 0,28 | | 0,56 | 0,2128 | 10,56 | 4,0128 | 4,0128 | 1,123584 | 20,5216 | 7,798208 | 7,798208 |
| 826 | 3 | 2,478 | 1 | 2,478 | 0,28 | | 0,56 | 0,69384 | 0,88 | 0,72688 | 0,72688 | 0,2035264 | 4,6464 | 3,8379264 | 3,8379264 |
| 1363 | 1 | 1,363 | 1 | 1,363 | 0,28 | | 0,56 | 0,38164 | 1,32 | 1,79916 | 1,79916 | 0,5037648 | 0,3872 | 0,5277536 | 0,5277536 |
| 1577 | 1 | 1,577 | 1 | 1,577 | 0,28 | | 0,56 | 0,44156 | 0,44 | 0,69388 | 0,69388 | 0,1942864 | 0,5808 | 0,9159216 | 0,9159216 |
| 4572 | 250 | 35,023 | 5,9 | 23,5359 | 0,28 | | 0,56 | 6,590052 | 222,56 | 33,0312 | 23,695476 | 6,63473328 | 210,7328 | 33,7770704 | 25,46159024 |
3) Для среднеэксплуатируемых популяций, у которых LC50 значительно больше LM50 (нет оснований для опасений за состояние запасов), применять правило F=M не рекомендуется, так как у эксплуатируемых популяций F всегда больше M.
Часть естественной смертности у особей промыслового запаса «скрыта» в промысловой смертности (то есть часть особей, которые неизбежно погибли бы от естественной смертности, вылавливаются промыслом). Считается, что для половозрелых особей в интенсивно облавливаемых популяциях от трети (для видов-жертв) до половины (для хищников, быстрорастущих и высокотелых видов рыб) естественной смертности может быть скрыто в промысловой. При этом, для видов рыб-жертв больше, для прочих видов - меньше. То есть, F больше M, так как часть М скрыта в F.
Для определения Z может быть использована таблица 27, где ранжирована Z в зависимости от предельного возраста T, либо F которая определена способом в соответствии с подпунктом 2), при этом F будет составлять 2/3 от Z для видов-жертв и 3/4 от Z для хищников, быстрорастущих и высокотелых видов рыб. Далее производится расчет ПДУ (пример расчета 2).
Таблица 27 Коэффициенты общей смертности Z (годовой убыли) и рекомендуемые коэффициенты изъятия F при достижении граничных ориентиров запаса, исходя из наблюдаемых значений предельного возраста рыб в уловах (выборке)
| Коэффициенты | Т (предельный возраст в уловах) |
| 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 |
| Z | 0,6 | 0,55 | 0,5 | 0,45 | 0,4 | 0,375 | 0,35 | 0,325 | 0,3 | 0,275 |
| F | 0,3 | 0,275 | 0,25 | 0,225 | 0,2 | 0,188 | 0,175 | 0,163 | 0,15 | 0,138 |
Пример расчета 2
| Возраст, лет | Средняя масса, грамм | Nt0, тысяч штук | Bt0, т | Половозрелость, доли единиц | Pt1, тонн | F | M | Z | ПДУ одного года, тонн |
| 1 | 35,9 | 55,2 | 2,0 | 0 | 0 | 0 | 0,35 | 0,35 | 0,0 |
| 2 | 91,7 | 111,4 | 10,2 | 0 | 0,0 | 0 | 0,35 | 0,35 | 0,0 |
| 3 | 232 | 328,6 | 76,2 | 0,5 | 38,1 | 0,131 | 0,219 | 0,35 | 5,0 |
| 4 | 493,1 | 324,8 | 160,1 | 1 | 160,1 | 0,262 | 0,088 | 0,35 | 42,0 |
| 5 | 746,1 | 80,0 | 59,7 | 1 | 59,7 | 0,262 | 0,088 | 0,35 | 15,6 |
| 6 | 1360,3 | 13,3 | 18,1 | 1 | 18,1 | 0,262 | 0,088 | 0,35 | 4,8 |
| 7 | 2200 | 8,6 | 18,9 | 1 | 18,9 | 0,262 | 0,088 | 0,35 | 4,9 |
| 8 | 3062,1 | 16,2 | 49,6 | 1 | 49,6 | 0,262 | 0,088 | 0,35 | 13,0 |
| 9 | 3932,5 | 2,9 | 11,2 | 1 | 11,2 | 0,262 | 0,088 | 0,35 | 2,9 |
| 10 | 6263,3 | 9,5 | 59,7 | 1 | 59,7 | 0,262 | 0,088 | 0,35 | 15,6 |
| 11 | 9060 | 1,9 | 17,3 | 1 | 17,3 | 0,262 | 0,088 | 0,35 | 4,5 |
| Всего | | 952,4 | 483,0 | | 394,5 | 0,262 | 0,088 | 0,35 | 108,4 |
Затем производится моделирование состояния запаса на 2 года вперед на соответствие критерию MSY аналогично, как в подпункте 2) пункта 112.
113. Биостатистический метод (далее - БСМ) оценки численности промыслового стада рыб в настоящее время применяется в отдельных случаях, когда оценка другими методами невозможна. Однако, в силу ряда причин, результаты от его применения оказываются заниженными. БСМ основан на возрастном распределении уловов с целью оценки численности поколений.
114. Во всех вариантах биостатистического метода важным этапом является составление расчетной таблицы, в которую сведены данные о ежегодных уловах исследуемого стада рыб в количественном выражении, распределенные по возрастным категориям.
115. Доля особей каждой возрастной группы получают из мониторинговых наблюдений (метод прямого учета) в период нерестовой миграции рыб (например, в реке Жайык). Сложением количества особей во всех поколениях, представленных в улове рассматриваемого года, находится минимальный (без учета естественной смертности) промысловый запас:
Vn = (1- х)Cn + (1- х - х1)Cn+1 + (1 - х - х1 - х2)Cn+2 +....+(1 - х - х1 - х2 - …. - х tλ)C tλ
где: n - запас рыбы в начале n-го года;
tλ - предельный возраст рыб;
Cn, Cn+1, Cn+2, ….. Ct
λ - уловы соответствующих лет;
х - процентное содержание в улове сеголеток;
х1 - процентное содержание в улове годовиков и так далее.
116. При определении предельно допустимых объемов процент изъятия высчитывается на основе «концепции репродуктивной разнокачественной популяции» Малкина по соответствующей методике (таблица 24).
117. Биостатистический метод оказывается приемлемым лишь при стабильности промысла, промысловых усилий и экологических условий.
118. Для вычисления нерестового запаса в таких случаях можно также учитывать кратность нереста и величину пополнения по возрастным группам, которую получают по данным летних морских траловых съемок:
NHI миллион экземпляров = NПI миллион экземпляров (остаток)*KН (кратность нереста) * Dcpi (доля созревающих рыб)
Таким образом, проводятся расчеты всех возрастных групп. Сумма всех поколений будет составлять нерестовый запас промысловых рыб. Биомасса нерестовой части популяции рассчитывается как произведение ее численности и средней навески особи, участвующей в нересте:

где: Bнз - нерестовый запас, тысяч тонн;
N - расчетная численность популяции, миллион экземпляров;
w - средняя масса особей, килограмм.
119. Таким же образом рассчитывают численность промыслового и нерестового запаса для последующих лет с учетом величины пополнения от особей, вступающих в промысел. Коэффициенты изъятия при определении предельно допустимых уловов высчитываются также на основе «концепции репродуктивной разнокачественной популяции» по соответствующей методике.
120. Основной целью методики Кушнаренко А.И. и Лугарева Е.С. является - оценка состояния популяции промысловых видов рыб, и разработка биологически обоснованного предельно допустимого объема их изъятия. За основу принята формула:
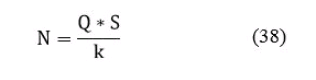
где: Q - количество рыб в контрольных уловах, в штуках; S - учетная площадь водоема в гектарах, получаемая вычленением непригодной для промысла зоны (заросли надводной растительности, большие глубины и так далее) из общей площади водоема; k - поправочный коэффициент, получаемый перемножением трех основных коэффициентов для каждой размерной группы (сети):
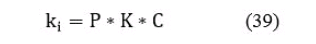
где: P - коэффициент вероятности встречи рыбы с орудиями лова; K - коэффициент уловистости сетей, C - площадь облова контрольного орудия лова. Данные коэффициенты вычисляются на основе экспериментальных данных.
121. Коэффициент Р вычисляется на основании формулы, указанной ниже. Коэффициент уловистости сетей К равен 0,2 - 0,5. Площадь облова определяется по формуле:
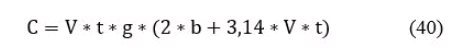
где: V - радиальная скорость рыскания, индивидуальная для вида (метр\минута); t - время сетепостановки в минутах; g - количество поставленных сетей; b - длина сети при стандартной высоте в 25 метров.
Где, основное значение имеет показатель V, значения которого определены в справочнике (Радаков Д.В. Протасов В.Р. «Скорости движения и некоторые особенности зрения рыб» - М.: Наука, 1964 - 48 страница).
122. Скорости рыскания для карася, окуня и щуки составляют 0,04, для леща и плотвы - 0,05, для карпа - 0,06, для линя - 0,10 и для судака - 0,13 метр/секунду.
123. В отдельных случаях, для ускоренной оценки численности рыб, допускается использование метода прямого подсчета гидроакустической съемкой современными рыбопоисковыми эхолотами с усовершенствованной функцией идентификации рыбы (FI) способных отличать рыб от других объектов, показать размеры рыбы и на какой глубине она зафиксирована.
124. При этом методе для прямого подсчета количества рыб разных размеров проводят эхолокацию акватории рыбопоисковым высокоэффективным эхолотом (модели марки Humminbird, Lowrance, Garmin и другие). Рекомендуемые характеристики эхолота следующие: минимум два луча сканирования, минимальный угол обзора - 20 градусов; глубина сканирования до 100 метров, минимальная частота сонара 200 килогерц; минимальная выходная пиковая мощность 800 ватт; минимальный размер экрана 7 дюймов, минимальный интервал между объектами не более 6,5 сантиметров, наличие датчиков температуры, скорости и пройденного расстояния, разбивка по размерам рыбы (мелкая, средняя, крупная), пиксельная матрица не менее 480Vx800H, тип цветной, подсветка LED, водонепроницаемость IPX7, возможность подключения к персональному компьютеру для последующей обработки и сохранения полученных данных.
125. Эхолокацию проводят путем передвижения с эхолотом по акватории на легкой весельной лодке со скоростью не более 3 километров/час (скорость определяют исходя из пройденного расстояния по показанию эхолота, и затраченного на это времени). При такой невысокой скорости результаты более достоверны. Схема эхолокации следующая: на водоемах площадью до 500 гектаров (рисунок 8, элемент 1) проводится сканирование от берега до берега (например, с северного на южный). От исходной точки на берегу (рисунок 8, элемент 2) маршрут сканирования (рисунок 8, элемент 3) идет к противоположной стороне водоема под углом 25 градусов. Достигнув берега, сканирование прекращают и проплывают вдоль берега и параллельно ему 100 метров. Затем от самого берега начинают сканирование в обратном направлении также под углом 25 градусов. И далее по такой схеме сканируется эхолотом вся акватория. Отклонение маршрута эхолокации на 25 градусов вызвано тем, чтобы расходящиеся под углом маршруты не захватывали уже зафиксированных рыб. Этой же цели служит и продвижение вдоль берега на 100 метров перед последующим сканированием. Учитывая невысокие радиальные скорости движения рыб, при такой схеме сканирования исключается повторное фиксирование рыб. По окончании первого сканирования, для сбора более полных данных, проводится второе сканирование, при котором маршруты от берега до берега отклоняются также на 25 градусов, но в другую сторону (рисунок 8, элемент 4), проходя по необследованным участкам. При наличии времени проводится еще один цикл сканирования водоема. Причем если в первом цикле маршруты проходили с северного берега на южный, то в втором цикле они должны проходить с восточного берега на западный. При большой площади водоема (от 1000 и более гектар) водоем разбивается на сектора с площадями 500 гектаров. При этом границы секторов и линии маршрутов сканирования предварительно фиксируются определением их географических координат спутниковым навигационным прибором типа Global Positioning System (далее - GPS), глобальной навигационной спутниковой системы (далее - ГЛОНАСС). Далее ведется сканирование эхолотом по аналогичной схеме. Замера длины маршрута сканирования проводится по данным эхолота. Если же в нем нет функции отображения пройденного расстояния, используется спутниковый навигационный прибор. В обязательном порядке исследуются выраженные участки биотопов - мелководье, глубина, омуты, эстуарий, тростниковая зона и другие.
Рисунок 8. Схема сканирования эхолотом водоема площадью до 500 гектаров
1 - сканируемая акватория;
2 - точка начала маршрута сканирования;
3 - маршрут первого цикла сканирования;
4 - маршрут второго цикла сканирования.
Эхолокацию водоема рекомендуется проводить в летнее время, когда нет миграций. Во время миграций рыба активно движется, что усложняет сканирование. В сутках, наиболее подходящее время для этой работы дневное - с 9 до 19 часов, когда основная масса рыб находится в состоянии покоя. В летнее время пик кормовой активности рыб приходится на ранее утро и поздний вечер.
Эхолоты последнего поколения показывают отдельных рыб с указанием размеров (номера от 1 до 8 и более, соответствующие разным размерам рыб, или в виде символов рыб различной величины). Если размеры рыб выражаются номерами, то они записываются в порядке, присущем используемой модели эхолота. Данные по размерам рыб каждого номера берутся из паспортных характеристик используемого эхолота. Если размеры рыб отображаются в виде символов различной величины, то этим символам необходимо присвоить соответствующие номера (мальки - 1, мелкая рыба - 2, средняя рыба - 3 и так далее. Данные по размерам рыб каждого вида символов, также берутся из паспортных характеристик используемого эхолота). Эхолот показывает, на какой глубине находится отдельная рыба. Эти данные записываются (таблица 28).
Таблица 28. Учет рыб в исследуемом водоеме по данным рыбопоискового эхолота
| Глубина, на которой зафиксирована рыба, метр | Количество разноразмерных рыб (в виде знака \), по номерам от 1 до 8 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
| 0,1 - 0,5 | \\\ | \ | \\ | \\\ | \ | \\ | - | \ |
| 0,5 - 1,0 | \\ | \\ | \\\ | \ | \ | \\ | \\ | - |
| 1,0 - 1,5 | \\\\ | \\ | \\\ | \ | \\\ | \ | \\\ | \ |
| и так далее | | | | | | | | |
| Примечание: если в используемой модели эхолота размеров рыб в цифровом или символьном отображении больше 8, то к таблице прибавляется аналогичное количество столбцов |
Глубина воды разбита на ряд слоев. Это обусловлено тем, что луч эхолота в 20 градусов, будет расширяться с удалением от поверхности воды, и в каждом последующем слое площадь сканирования будет последовательно увеличиваться. Определение численности рыб по данным сканирования эхолотом проводится отдельно для каждой размерной группы и каждого отдельного слоя воды толщиной 0,5 метра. Первоначально необходимо подсчитать количество разноразмерных рыб, зафиксированных в площади сканирования.
Подсчет рыб на глубине до 0,5 метров отдельно по каждому размеру.
Ширина полосы сканирования первого слоя воды до глубины 0,5 метров является расчетной шириной для определения общей площади сканирования. В последующих слоях ширина полосы сканирования (с расширением угла луча) увеличивается, поэтому результаты расчетов количества рыб по всем последующим слоям, пересчитываются на ширину полосы первого слоя.
Подсчет количества рыб в этом, и всех последующих слоях, ведется отдельно по каждому размеру рыб. При сканировании закрепленный на лодке эхолот (рисунок 9, элемент 5) расположен в 20 сантиметрах выше поверхности воды (элемент 6). Луч эхолота (элемент 7), направленный вертикально вниз в воду, имеет угол 20 градусов.
Рисунок 9. Луч эхолота при сканировании, в разрезе вид сбоку
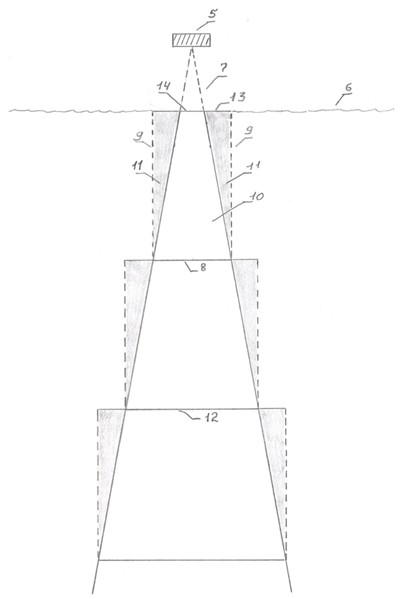
5. Эхолот;
6. Поверхность воды;
7. Вершина луча от эхолота до поверхности воды;
8. Основания луча в первом слое воды;
9. Вертикальные линии от основания луча до верхней границы слоя;
10. Охваченный сканированием участок;
11. Неохваченные сканированием участки;
12. Основание луча во втором слое;
13. Катеты одного из прямоугольных треугольников - неохваченных сканированием участков;
14. Основание луча от эхолота до поверхности воды.
При достижении глубины 0,5 метров основание луча (элемент 8) составит 24,6 сантиметра, рассчитывается по следующей формуле:

где: с - длина основания, а - длина перпендикуляра от вершины луча до центра основания и равная 50 сантиметрам, tg 10 - тангенс половины угла луча дающая значение 0,176.
Для расчета площади сканирования в этом слое, от обеих крайних точек основания луча, проводится до верхней границы слоя (поверхность воды), две вертикальные линии (элемент 9). Образуется четырехугольный прямоугольник шириной 24,6 сантиметров и высотой 50 сантиметров, который обозначается как wb (Примечание: данная ширина принимается за ширину отсканированной полосы, которая при умножении на длину маршрута, дает величину площади сканирования). Однако в прямоугольнике wb имеется охваченный сканированием участок w (элемент 10) и неохваченный b (элемент 11) в соотношении 64,3 % на 35,7 %. Это соотношение вычисляется следующим образом: площадь wb равна 1230 см2; площадь b состоит из двух аналогичных прямоугольных треугольников. Определяется площадь одного из них, которая будет равна произведению катетов, разделенного на 2. Поскольку один из катетов - это высота слоя воды, то его длина составит 50 сантиметров. Длина второго катета (элемент 13) составит 8,78 сантиметра, что равно половине разности длины основания луча в слое (рисунок 9, элемент 8) и длины основания луча от эхолота до поверхности воды (элемент 14) составляющая 7,04 сантиметра.
Примечание: во втором и далее слоях, при этом расчете берутся длина основания луча в слое и длина основания луча в верхнем слое.
Тогда площадь одного треугольника будет равна 219,5 сантиметр2. Площадь b определяется, как сумма площадей этих двух треугольников, она равна 439 сантиметр 2. Тогда площадь w будет равна разности площадей wb и b, и составит 791 сантиметр 2. Далее площади wb, w и b переводятся в проценты. При этом wb будет равно 100 %, w - 64,3 %, b - 35,7 %.
Исходя из количества рыб f в секторе w, полученных из данных эхолота рассчитывается количество рыб в секторе b по следующей формуле:
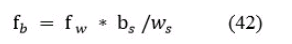
где: fb - количество рыб одного размера в секторе b, штук;
fw - количество рыб одного размера в секторе w, штук;
bs - площадь сектора b, сантиметр2;
ws - площадь сектора w, сантиметр2.
Общее количества рыбы одного размера в этом слое составит:

где: f - количество рыб одного размера, штук;
fw - количество рыб в секторе w, штук;
fb - количество рыб в секторе b, штук.
Учитывая, что эхолот показывает рыбу по размерам (от 1 до 8), то f определяется для каждого размера.
Подсчет рыб на глубине до 0,5 - 1,0 метра ведется аналогично. Однако в этом втором слое воды при достижении глубины 1,0 метра основание луча составит уже 42,24 сантиметра. Следовательно, площадь сканирования во втором слое будет больше расчетного. Поэтому количество рыб в данном слое необходимо привести к расчетной площади сканирования, следующим образом:
расчетная площадь сканирования первого слоя составляет 58 % от площади сканирования второго слоя. Поэтому во втором слое мы учитываем только 58 % от общего количества рыб. Подсчеты рыб на всех последующих глубинах проводят аналогично.
Подсчет количества рыб различных размеров в площади полосы сканирования.
Площадь полосы сканирования определяется по формуле:

где: С - площадь полосы сканирования эхолотом, метр2;
L - длина маршрута сканирования, метр;
a - расчетная ширина полосы сканирования в метрах, равная при луче с углом 20 градусов 0,246 метра.
Количество рыб каждого размера в площади сканирования, определяется по следующей формуле:

где: n - количество рыб каждого отдельного размера в площади С;
∑f - сумма количеств рыбы отдельного размера в слоях.
Общее количество рыб каждого размера на площади водоема, определяются по формуле:
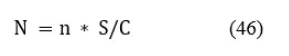
где: N - общее количество рыб отдельного размера на площади водоема S;
n - количество рыб этого размера в площади сканирования С;
S/C - отношение площади водоема к площади сканирования.
Значение N рассчитывается отдельно по всем размерным группам, зафиксированным при сканировании эхолотом.
После определения количества рыб каждого размера на водоеме, необходимо определить данные по видовому, размерному, весовому составу рыб. Для этого проводятся ловы научно-исследовательскими орудиями лова с размерами конструктивного шага ячеи от 20 до 80 миллиметров. Изучение биологических характеристик проводится по общепринятым в ихтиологии методикам. По данным вылова устанавливается видовой, размерный и весовой состав промысловой ихтиофауны, по размерным группам 1-8.
Например, в размерной группе 3 присутствуют: сазан 15 %, судак 10 %, лещ 75 %. Общее количество рыб размерной группы 3 (по данным эхолокации) - 300 штук. Следовательно, в водоеме будет 45 экземпляров сазана, 30 экземпляров судака, 225 экземпляров лещ, данной размерной группы. Таким же образом по всем размерным группам, которые выдаются эхолотом.
Далее, путем взвешивания рыб из уловов определяется средняя масса рыбы каждого вида, в каждой размерной группе. Средняя масса умножается на количество рыб (при этом учитываются только рыбы, достигшие промысловых размеров).
Таким образом, получается биомасса рыбы одного вида в отдельных размерных группах 1, 2, 3….8. Данные значения суммируются и получается общая биомасса рыбы этого вида в водоеме, то есть промысловый запас. Полученная величина промыслового запаса рыб по видам, является основой для расчета общего допустимого улова.
126. Все методы учета численности рыб (метод площадей, методика Кушнаренко-Лугарева, метод Мельниковой, методика ВНИПРХ) имеют значительную погрешность, так как включают трудноопределимые коэффициенты уловистости применяемого орудия лова.
В водоемах республиканского и международного значения оценка численности, для проверки адекватности и достоверности результатов определения численности рыб, проводится 2 методами - основным (вышеупомянутые методики) и вспомогательным (упрощенный метод эхолотно-сетной съемки, позволяющий упростить и ускорить сбор и обработку материалов гидроакустической съемки), что позволит скорректировать в основном методе коэффициенты уловистости применяемых орудий лова в зависимости от полученных значений численности рыб при эхолотно-сетной съемке («коэффициент уловистости» эхолота близок к 1). Сущность метода изложена в примере ниже.
Пример. Определение численности рыб в Верхне-Тобольском водохранилище.
Проведена эхолотно-сетная съемка (6 эхолотных разрезов и 3 сетепостановки). Каждая эхолотно-сетная съёмка выполняется в три этапа. На первом этапе по месту предполагаемой постановки сетей осуществлялся проход на лодке с эхолотом для учета численности рыб на данном участке водоёма. Для упрощения расчетов параметры охвата луча определяются для глубины, на которой зафиксированы наибольшие скопления рыбы, затем диаметр луча эхолота на этой глубине умножается на расстояние, пройденное при эхолотировании и находится площадь съемки. На втором этапе осуществлялась постановка стандартного порядка сетей (ячея от 20 до 70 миллиметров, 9 сетей по 25 метров), продолжительность постановки сетей составляла 300 минут (с 11.00 до 16.00). На третьем этапе по месту постановки сетей осуществлялся проход на лодке с эхолотом для учета численности рыб на данном участке водоёма (таблица 29).
Таблица 29. Сведения по количеству рыб, учтенных при каждом эхолотировании
| Эхолотный разрез | Количество рыб, экземпляр |
| 1 | 98 |
| 2 | 77 |
| 3 | 56 |
| 4 | 84 |
| 5 | 35 |
| 6 | 56 |
| Всего рыб, экземпляр | 406 |
Длина эхолотного разреза 250 метров при глубине 3 метра, таким образом, площадь, пройденная эхолотом, составляет 250 метров*3,464 метров = 866 метров2. Всего было сделано 6 эхолотных разрезов, таким образом, просканирована площадь равная 866 метров2*6 = 5196 метров2.
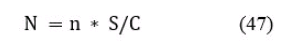
где: N - общее количество рыб на площади водоема S;
n - количество рыб на площади сканирования С;
S/C - отношение площади водоема к площади сканирования.
По формуле находится общее количество рыб в Верхне-Тобольском водохранилище: N = 406 экзепляров* 87400000 метров2/5196 метров2= 6829176 экземпляров. По результатам сетепостановок определяем процентный состав ихтиофауны: плотва - 20,5 %, карп - 1,5 %, лещ - 52,8 %, щука - 0,9 %, окунь - 19,3 %, судак - 1,8 %, рипус - 3,2 % (таблица 30).
Таблица 30. Разбивка по видам рыб (%)
| |
| плотва | карп | лещ | щука | окунь | судак | рипус |
| 20,5 | 1,5 | 52,8 | 0,9 | 19,3 | 1,8 | 3,2 |
Исходя из полученных данных, рассчитывается численность каждого вида (таблица 31).
Таблица 31. Расчет численности рыб по видам
| Вид | Общая численность | Промысловая часть популяции | Промысловая численность | Средняя навеска, килограмм | Промысловый запас, тонна |
| плотва | 1399981 | 0,63 | 881988 | 0,1 | 88,20 |
| карп | 102437 | 0,31 | 31755 | 1,32 | 41,92 |
| лещ | 3605805 | 0,56 | 2019251 | 0,17 | 343,27 |
| щука | 61462 | 0,38 | 23356 | 1,45 | 33,87 |
| окунь | 1318031 | 0,67 | 883081 | 0,11 | 97,14 |
| судак | 122925 | 0,22 | 27044 | 1,39 | 37,59 |
| рипус | 218533 | 1 | 218533 | 0,09 | 19,67 |
127. Сбор биологических показателей рыб при прижизненном исследовании включает в себя измерение длины тела рыбы, взвешивание, отбор регистрирующих структур на определение возраста рыб (в общем случае - чешуя, у осетровых видов рыб - часть первого луча грудного плавника). В лаборатории определяется возраст рыб, рассчитывается коэффициент упитанности по Фультону.
128. Оценка численности редких и находящихся под угрозой исчезновения видов рыб такая же, как и промысловых видов.
Однако, в отдельных случаях, когда производители рыб проходят на нерест в мелководные неширокие речки и ручьи с прозрачной водой, может применяться визуальный подсчет проходящих производителей и определение численности повременным методом, фото- и видеосъемка («прижизненным методом»).
129. При проведении исследований совместно с зарубежными учеными на трансграничных водоемах, могут использоваться иные методики, если это оговорено международными соглашениями.
130. В отдельных случаях, иные методики исследований могут использоваться на водоемах и (или) участках специального назначения, например, в охраняемой зоне верхнего и нижнего бьефов гидроэлектростанции (ГЭС), где отдельные исследования традиционными методами затруднены. Методика таких работ должна быть тщательно обоснована в отчете. Во всех остальных случаях осуществляется применение вышеописанной «единой методики».
131. Работы по определению запасов цист артемии включают в себя:
1) Определение географического положения водоема;
2) Изучение метеорологических условий в период сбора материала;
3) Изучение морфометрических характеристик и исследование гидрологического режима водоемов;
4) Изучение гидрохимического режима водоемов;
5) Проведение гидробиологических исследований;
6) Расчеты запасов цист артемии и определение общих допустимых уловов.
132. Определение количества и места расположения станций по отбору проб на водоемах проводится согласно методическим рекомендациям по сбору и обработке материалов при гидробиологических исследованиях. Координаты станций определяются с помощью навигационной системы GPS или ГЛОНАСС.
133. Сроки отбора проб должны быть связаны с жизненными циклами и изменениями условий обитания исследуемого объекта. Для рыбных ресурсов прогноз ПДУ выполняется с учетом всех сезонов года. Для определения запасов цист и подготовки окончательного прогноза ПДУ научно-исследовательские работы должны выполняться в вегетационный период.
134. В отчете в обязательном порядке приводятся географические сведения о водоеме (географические координаты, месторасположение с привязкой к населенным пунктам). Географические координаты определяются с помощью навигационной системы GPS или ГЛОНАСС. Месторасположение с привязкой к населенному пункту определяется кратчайшим расстоянием от границы населенного пункта до береговой линии водоема. Сведения приводятся в виде таблицы (таблица 32).
Таблица 32. Координаты и месторасположение водоемов
| Водоем | Район | Место расположения | Координаты |
135. Метеорологические исследования необходимо проводить на каждой станции отбора проб. В метеорологический журнал заносятся сведения о силе и направлении ветра, температуре воздуха, интенсивности солнечного света и силе волн с указанием даты, времени и координат определения метеорологических параметров. Часть метеорологических параметров определяется визуально, часть - с помощью специальных приборов.
136. Изучение морфометрических характеристик и гидрологического режима водоемов включает в себя определение следующих параметров:
характер водосборной площади, наличие и состояние притоков;
площадь водного зеркала;
длина и наибольшая ширина водоема;
длина береговой линии;
развитие береговой линии и наличие заливов;
максимальная и средняя глубина;
объем водной массы;
изменение уровня воды по сезонам и по годам (для водоемов, где осуществляется ежегодный мониторинг).
137. Изучение водосборной площади проводится путем визуальных наблюдений. Площадь акватории, длина береговой линии, длина и наибольшая ширина водоема определяется путем измерения на местности с помощью навигационных систем или с использованием спутниковых снимков. Промеры глубин проводятся с помощью лота, либо эхолота. Уровень воды определяется с помощью навигационной системы. По результатам изучения морфометрических исследований и определения максимальных и средних глубин специалистами производится определение «жилой» зоны для артемии с учетом биологических особенностей этого вида, гидрологического режима водоема и гидрометеорологических условий. Полученные результаты отражаются в виде таблицы (таблицы 33, 34).
